
Termin klonowanie pochodzi od greckiego słowa klon, czyli pęd, latorośl, gałązka. Do naukowego słownika wszedł on na początku XX wieku i oznaczał grupę roślin propagowanych przy użyciu jakichkolwiek części wegetatywnych. Dziś tym mianem określa się proces tworzenia dokładnych kopii organizmu macierzystego, czyli osobników zawierających identyczną informację genetyczną klonów.
Pierwowzorem klonowania jest proces rozmnażania wegetatywnego, stanowiącego jedną z form bezpłciowej reprodukcji. Potomstwo powstaje na drodze bezpośrednich mitotycznych podziałów komórkowych i uzyskuje zestaw genów identyczny z informacją zawartą w komórkach organizmu rodzicielskiego. Proces ten umożliwia szybkie zasiedlenie danej niszy ekologicznej, jednak brak zmienności staje się przyczyną gorszego dostosowania do panujących warunków środowiskowych. Ten typ rozmnażania występuje powszechnie u glonów, bakterii, pierwotniaków oraz roślin (poprzez kłącza, rozłogi, cebule, bulwy i in.) i jest często wykorzystywany w ogrodnictwie przy uprawie odmian o pożądanych cechach użytkowych.
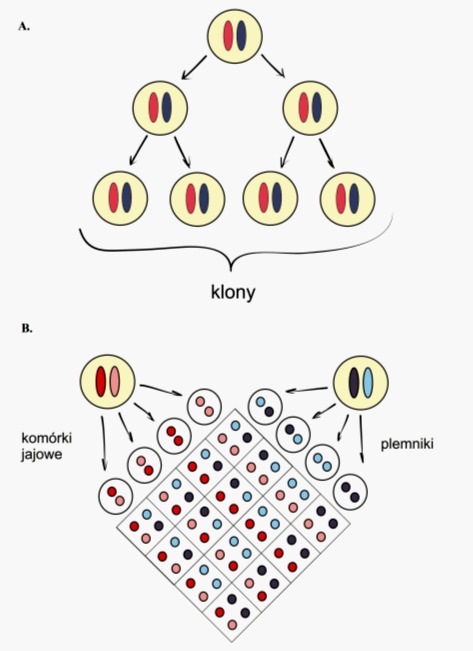
Rys. 1. W wyniku klonowania, tak jak podczas rozmnażania wegetatywnego, uzyskuje się osobniki identyczne pod względem genetycznym (A). Dzięki rozmnażaniu płciowemu następuje tzw. zmienność genetyczna (B)
Klonowaniem jest również dzieworództwo, czyli partenogeneza, podczas której organizm potomny kształtuje się z niezapłodnionej komórki jajowej. Brak udziału plemnika umożliwia dokładne skopiowanie materiału genetycznego. Zjawisko to występuje powszechnie w świecie zwierząt, np. u owadów. W warunkach naturalnych nie obserwuje się go jednak u ssaków.
Klonami są także bliźnięta jednojajowe, czyli dwa płody rozwijające się z jednej zapłodnionej komórki jajowej. Normalnie, węzeł zarodkowy zawiązuje się jako pojedyncza grupa komórek. W przypadku ciąży bliźniaczej tworzą się dwie takie grupy. Potomstwo rozwija się zatem z jednej pary gamet i dlatego posiada identyczny materiał genetyczny. Zjawisko to występuje dość rzadko. U koni ciąża bliźniacza zostaje zazwyczaj niedonoszona, gdyż łożysko nie jest w stanie odżywić obu rozwijających się osobników. Istnieją także wyjątki. Ciąże bliźniacze są regularne u pancerników, których zarodki mogą się dzielić nawet na kilkanaście części.
Pierwsze eksperymenty związane z klonowaniem wynikały z czystej, naukowej ciekawości. Od lat pięćdziesiątych przeprowadzano próby z klonowaniem żab, których skrzek jest łatwo dostępny i przezroczysty. Takie manipulacje były możliwe bez udziału zaawansowanej technologii. Dzięki embriologii eksperymentalnej wykazano, że po wyjęciu jądra z komórki zarodkowej i wprowadzeniu go do pozbawionej jądra komórki jajowej, zaszczepiony materiał potrafi pokierować całym dalszym rozwojem. Nie udało się jednak uzyskać podobnych wyników w przypadku pobrania materiału genetycznego z już rozwiniętych komórek, np. pochodzących z jelita, gdyż niektóre ich geny były na stałe zablokowane. Próby nie powiodły się również w przypadku myszy, u których geny zaczynały funkcjonować już na etapie dwukomórkowego zarodka. Najbardziej korzystne okazały się zwierzęta gospodarskie, które mimo swoich dużych rozmiarów i nieprzezroczystej komórki jajowej zapewniały odpowiedni czas na przeprowadzenia manipulacji genetycznych.
Pierwszym sklonowanym ssakiem, pochodzącym z komórek dorosłego osobnika była owieczka Dolly. Dokonał tego zespół dr. Iana Wilmuta z ośrodka Agriculture Research Centre w Cambridge. Do eksperymentu posłużono się czterema owcami nazwanymi dziś matkami Dolly. Pierwsza z nich była dawczynią oocytu, z którego następnie usunięto jądro i zastąpiono je informacją genetyczną, pochodzącą z somatycznych komórek gruczołu mlecznego drugiej owcy. Wilmut opracował specjalną metodę opartą na głodzeniu hodowanych komórek, tzn. zastosowaniu obniżonej zawartości surowicy w pożywce. Spowodowało to, że komórki somatyczne weszły z powrotem w fazę cyklu G0, co znacznie ułatwiło ich przeprogramowanie. Fuzji z oocytem dokonywano za pomocą prądu elektrycznego. Łącznie przeprowadzono 277 takich prób i uzyskano 247 połączonych zarodków, które następnie przeniesiono do podwiązanych jajowodów owiec – biorczyń pośrednich (numer 3). Ich rolą było zapewnienie dogodnych warunków rozwoju, niemożliwych do uzyskania w hodowli in vitro. 29 zarodków przetransplantowano następnie do 13 owiec – biorczyń ostatecznych (numer 4). U jednej z nich stwierdzono ciążę i w ten sposób 5 lipca 1996 roku narodziła się owieczka Dolly.
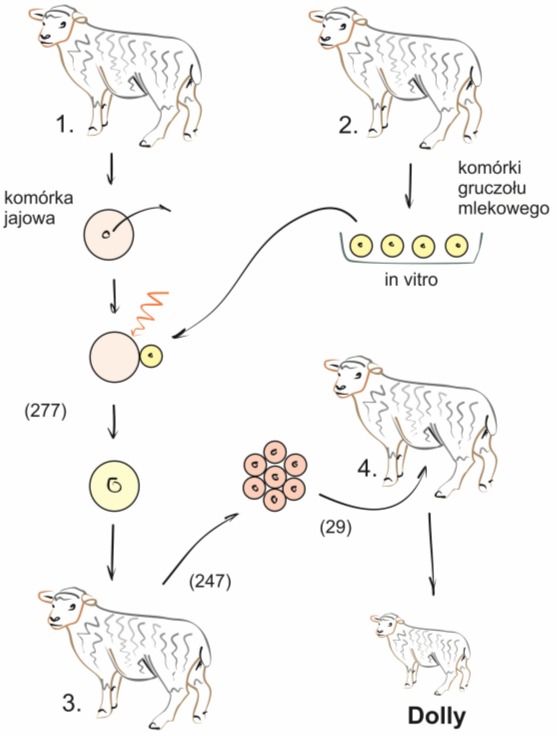
Rys. 2. Uproszczony schemat procedury transferu jadra komórkowego, dzięki któremu powstała owieczka Dolly
Od tego czasu technika transferu jąder komórek somatycznych (ang. somatic cell nuclear transfer – SCNT) pozwoliła sklonować wiele różnych gatunków ssaków: myszy, bydła, kozła, świni, wołu, muflona, kota domowego, królika, konia, muła, szczura, kota nubijskiego, psa, fretki, wilka, jelenia, kota arabskiego, koziorożca alpejskiego, wielbłąda.
Metoda ta ma ogromny potencjał, jednak u niektórych gatunków obserwuje się bardzo niski procent skuteczności i wysoką śmiertelność u uzyskiwanego potomstwa. Przykładowo, skuteczność u myszy wynosi jedynie 1-2%. Komplikacje wynikają głównie z niekompletnego przeprogramowania się jąder komórek somatycznych. Opracowywano wiele strategii mających na celu wzmocnienia efektywności metody SCNT:
• wyrównanie etapów cyklu rozwojowego komórek dawcy, a także synchronizacja pomiędzy komórkami dawcy i oocytami,
• stosowanie komórek somatycznych od dawców w różnym wieku, pochodzących z różnych tkanek, pasaży oraz hodowanych w różnych warunkach,
• transfer komórek macierzystych o niskim poziomie znaków epigenetycznych,
• modyfikacja znaków epigenetycznych komórek dawcy za pomocą leków.
Większość z modyfikacji skupia się na komórkach somatycznych. Żadna z wymienionych metod nie zdołała jednak przezwyciężyć problemów związanych z etapem transferu jądra do oocytu.
Dzięki eksperymentom Wilmuta dowiedzieliśmy się, że raz zróżnicowana komórka zwierzęca nie ma w nieodwracalny sposób zablokowanej informacji genetycznej. Od czasu owieczki Dolly narodziło się jednak wiele pytań odnośnie znaczenia i etyki klonowania zwierząt.
Sklonowane embriony wykazują rozwojowe anomalie, takie jak np. nadmierny rozrost, czyli tzw. syndrom dużego potomstwa (ang. large-offspring syndrome – LOS). Wiele organizmów jest również dotkniętych otyłością oraz wrodzonymi defektami wątroby i układu immunologicznego. Ich komórki posiadają telomery o zmienionej długości, odziedziczonej najprawdopodobniej po dorosłej komórce dawcy. Zmniejsza to w znaczący sposób żywotność klonów.
Sztuczne populacje powielonych osobników nie są również zadowalające z punktu widzenia ewolucji. Przypominają one unikaną przez człowieka hodowlę w pokrewieństwie. Cenne może być jedynie kopiowanie jednostkowych osobników o wybitnie korzystnych cechach, np. koni wyścigowych. Klonowania nie można również wykorzystać do ratowania zagrożonych gatunków, gdyż groziłoby to wystąpieniem jednolitości genetycznej. Można za to powielać zwierzęta transgeniczne, produkujące cenne substancje lub farmaceutyki. Przeprowadzona transformacja może bowiem uniemożliwić rozród płciowy, a ewentualne, naturalne potomstwo może zostać pozbawione korzystnych cech.
Istnieje również klonowanie na poziomie zarodkowym, polegające na uzyskiwaniu bliźniąt w wyniku podziału zarodków. Ma to wieloletnią tradycję i dotyczy głównie bydła. Klonując zarodek wiemy, że ma on wartościowych rodziców, nie możemy być jednak pewni, że potomek odziedziczy wszystkie pożądane cechy.
Technika klonowania może być także stosowana do pozyskiwania genetycznie dopasowanych zarodkowych komórek macierzystych, wykorzystywanych do celów terapeutycznych. Sklonowane embriony nie byłyby umieszczane w macicy, a jedynie stanowiłyby potencjalne źródło totipotencjalnych komórek, z których w razie potrzeby można by było wyhodować tkanki i organy do przeszczepów.
Idea klonowania człowieka stanowi jednak kontrowersyjne zagadnienie i wywołuje wiele dyskusji na tle etycznym.
Autor: Anna Kurcek
Literatura:
• Ian Wilmut, Keith Campbell, Colin Tudge „Ponowny akt stworzenia. Dolly i era panowania nad biologią”, Dom Wydawniczy Rebis, Poznań 2002;
• Damian Strączek „Klonowanie”, Wydawnictwo Znak, Kraków 1998;
• Nguyen Van Thauan, Stoshi Kishigami, Teruiko /Nakayama „How to Improve the Success Rate of Mouse Cloning Technology”, Journal of Reproduction and Development, Vol. 56, No. 1, 2010;
• Cindy Tian, Chikara Kubota, Brian Enright, Xiangzhong Yang “Cloning animals by somatic cell nuclear transfer – biological factors”, Reprod Biol Endocrinol. 2003; 1: 98;
• Anna-Katerina Hadjantonakis, Virginia E Papaioannou “Can mammalian cloning combined with embryonic stem cell technologies be used to treat human diseases?”, Genome Biol. 2002; 3(8): reviews1023.1–reviews1023.6;
• Jie Xu, Xiangzhong Yang “Will cloned animals suffer premature aging – The story at the end of clones’ chromosomes”, Reprod Biol Endocrinol. 2003; 1: 105;