
Autor: Paweł Krzyżek
Endofity to organizmy, które kolonizują tkanki gospodarza nie wywołując przy tym objawów chorobowych. Każdy gatunek rośliny jest gospodarzem dla wielu endofitów, jednak zazwyczaj jeden lub dwa są dominujące. W naturze rośliny nie są w stanie bez nich przeżyć.
Kolonizacja rośliny przez endofity zachodzi przez różne wrota. Źródłem endofitycznych bakterii mogą być nasiona, zwłaszcza te, które zostały w jakikolwiek sposób uszkodzone. Jeśli doszło do wdarcia się przez okrywy nasienne, endofity takie będą uprzywilejowane. Fylosfera, czyli powierzchnia rośliny wystająca ponad glebę, to kolejne miejsce wnikania drobnoustrojów. Wrotami przeniknięcia przez fylosferę są aparaty szparkowe, przetchlinki i wszystkie uszkodzenia mechaniczne. Ryzosfera, powierzchnia podziemna, stanowi jednak główne miejsce interakcji między drobnoustrojami glebowymi oraz roślinami i to właśnie tu znajduje się największa liczba endofitów. Wynika to z łatwiejszej możliwości przeniknięcia przez cienką, nierzadko silnie uszkodzoną epidermę z licznymi włośnikami. Powierzchnia fylosfery jest mniej przyjazna dla mikroorganizmów ze względu na duże wahania temperatur oraz małą dostępność wody i substancji odżywczych.
Na kolonizację rośliny składa się prekolonizacja, czyli całość procesów dążących do wniknięcia do wnętrza rośliny i postkolonizacja, której celem jest namnożenie oraz odpowiednia lokalizacja w tkankach.
Wydzieliny ryzosfery stanowią bodziec dla bakterii endofitycznych które migrują w kierunku jego źródła w procesie chemotaksji. Jednym z najistotniejszych czynników komunikacyjnych w tym procesie są flawonoidy.
Następny etap stanowi adhezja do powierzchni korzenia. Początkowo są to słabe, odwracalne oddziaływania międzycząsteczkowe, które potem przekształcone zostają w nieodwracalne wywołane wiązaniem przy pomocy fimbrii, flagelli lub otoczek. Bakterie znajdujące się w fazie logarytmicznego wzrostu silniej adherują niż w fazie stacjonarnej.
Po procesie specyficznego rozpoznania roślina-endofit dochodzi do penetracji korzeni przez bakterie. Drogi zasiedlenia stanowią naturalne otwory w postaci porów i mikroporów, a także wszelkiego rodzaju uszkodzenia oraz rany. Endofityczne bakterie mają także zdolność do wytwarzania enzymów celulolitycznych i proteolitycznych, umożliwiających wniknięcie przez powłoki roślinne. Różnice między endofitami a patogenami są takie, że synteza u tych pierwszych odbywa się na niskim poziomie, natomiast u bakterii patogennych ekspresja takich enzymów jest wysoka. Drugą cechą odróżniającą jest zatrzymanie produkcji enzymów litycznych przez endofity w momencie skolonizowania przestrzeni międzykomórkowych, zaś u patogenów ich produkcja jest stała.
Rys. 1. Zdjęcie z mikroskopu fluorescencyjnego, obrazujące ryzosferę z otaczającymi ją wybarwionymi na czerwono filamentami grzybów i komórkami bakterii endofitycznych (źródło: http://www.personal.psu.edu).
Oddziaływanie endofitów na rośliny może być pozytywne, negatywne lub neutralne. Najbardziej interesujący z punktu promocji rozwoju roślin, jest aspekt pozytywny. Większość korzyści osiąganych jest przez produkcję metabolitów. Substancje te odgrywają rolę nie tylko w ochronie gospodarza i kompetycji z patogenami, ale również w specyficznych interakcjach i komunikacji roślina-endofit. Główne znacznie mają tu metabolity wtórne. Nie posiadają żadnych specyficznych funkcji w organizmie ich producenta, natomiast wywierają ogromny wpływ na interakcje międzyorganizmalne w ich otoczeniu.
Endofity przystosowały się do specyficznych warunków panujących wewnątrz organizmu rośliny i okazuje się, że ich potencjał metaboliczny różni się od tych, znajdujących się w glebie. Obligatoryjne endofity posiadają mniej bogaty arsenał metabolitów, mających znaczenie antybakteryjne i antypasożytnicze, ale ich siła metaboliczna skupia się na zwiększeniu panelu metabolitów odpowiadających za interakcje z gospodarzem. Życie w interakcji z roślinami wymaga adaptacji do wielu nisz, co przekłada się na zwiększoną ilość metabolitów wtórnych. Z drugiej strony, endofity wyspecjalizowane w interakcji z jednym określonym gatunkiem roślin i kolonizujące jedynie ściśle określone nisze produkują potencjalnie mniej metabolitów wtórnych. Jednakże wiele bakterii to endofity fakultatywne, radzące sobie dobrze zarówno w ryzosferze, jak i we wnętrzu rośliny.
Fitohormony odgrywają ogromną rolę w rozwoju roślin. Modulowanie ilości wytwarzanych hormonów przez endofity może mieć ogromne znaczenie w roślinnym sygnalingu. Bakterie te mają zdolność do produkcji kwasu indolilo-3-octowego (IAA), cytokinin i giberelin, co powoduje bardziej rozłożysty wzrost korzeni. Prócz tego posiadają także zdolność modyfikacji poziomu etylenu, który odpowiada za dojrzewanie i procesy starzenia. Drobnoustroje syntezują enzym deaminazę ACC, ta zmniejsza produkcję etylenu, a tym samym powstrzymuje starzenie rośliny i stymuluje rozrost korzeni. Roślina uzyskuje dzięki temu większą powierzchnię chłonną, a endofity zwiększoną ilość wrót wniknięcia do gospodarza. Co ciekawe, bakterie są w stanie produkować nie tylko fitohormony, ale również cząsteczki o strukturalnej analogii, pełniące podobną funkcję.
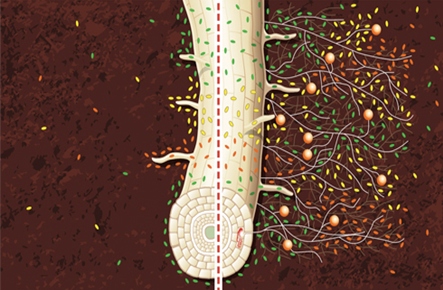
Rys. 2. Obrazek ukazujący zależność między rozgałęzieniem korzeni rośliny a ilością endofitów w ryzosferze (źródło: http://www.ganeshtree.com).
Występowanie bakterii w przestrzeniach międzykomórkowych powoduje odsunięcie się od siebie komórek roślinnych i wzrost powierzchni chłonnej, przez którą mogą pobierać substancje pokarmowe. Co więcej, endofity mają zdolność do mineralizacji związków organicznych siarki i fosforu i udostępniania go roślinom.
Jedną z korzyści współżycia z bakteryjnymi endofitami jest wiązanie wolnego azotu do form przyswajalnych dla roślin, co tłumaczy możliwość wzrostu na glebach ubogich w ten pierwiastek. Organizmy endofityczne zawierają geny umożliwiające im ekspresję enzymu, nitrogenazy, który odpowiada za redukcję wolnego azotu do amoniaku. W zamian za udostępnianie roślinom azotu, endofity zostają obficie zaopatrzone w potrzebne im substancje odżywcze oraz uzyskują protekcję przed nadmiernym stężeniem tlenu, wpływającym negatywnie na wiązanie azotu.
Interesującym jest fakt, że obecność endofitów w tkankach roślin promuje intensywniejszą infekcję grzybami mikoryzowymi, a przez to wzrost dostępności fosforu dla gospodarza.
Asocjacja roślin z endofitami warunkuje kontrolę biologiczną, czyli indukuje oporność gospodarza na patogeny. Wyróżnia się dwa rodzaje: działanie antagonistyczne oraz indukcję systemicznej oporności (ISR). Antagonizm wywołuje jednorazowy skutek i celuje w wąskie spektrum gatunkowe, natomiast ISR wykorzystuje kaskadę mechanizmów obronnych rośliny, które są jedynie aktywowane przez bakterie endofityczne, a zakres działania jest szeroki.
Do antagonizmu zaliczana jest produkcja antybiotyków oraz sideroforów. Siderofory posiadają duże powinowactwo do żelaza, odbierając je mikroorganizmom patogennym. Zmniejszenie stężenia żelaza, ważnego makroelementu dla bakterii, ogranicza ich wzrost, a tym samym promuje rozwój roślin. Prócz tego siderofory jako chelatory żelaza wiążą jony Fe3+. Taki kompleks jest kierowany do protoplastu roślin i w przestrzeni peryplazmatycznej zostaje zredukowany do jonów Fe2+, a te mogą być już wykorzystane w metabolizmie komórek roślinnych. Siderofory mają także zdolność do wiązania innych pierwiastków jak glin, miedź, kadm czy gal, a te zaopatrują gospodarza.
Przykładem systemicznej oporności jest wytwarzanie przez endofity biopolimerów, które przyczyniają się do lignifikacji ścian komórkowych roślin, wzmacniając bariery ochronne.
Endofity mają również spore znacznie w fitorekultywacji, a więc usuwaniu ze środowiska ksenobiotyków oraz powodują wzrost tolerancji rośliny na metale ciężkie np. przez tworzenie nierozpuszczalnych soli kwasów organicznych.
Większość bakterii nie daje się hodować w laboratoriach, co powoduje, że wiedza na temat relacji gospodarz roślina-endofit jest niewielka. Współzależność przynosi ogromne zyski dla obu stron. Bakteryjne endofity oddziałują na nadziemne i podziemne tkanki roślinne promując wzmożony rozwój roślin, zarówno w warunkach normalnych, jak i stresowych. Endofity mają zdolność intensywnych podziałów w miejscach uszkodzeń i ataku patogennych organizmów, przez co w wyniku konkurencji usuwają je z tych nisz. Liczebność endofitów jest stale monitorowana i regulowana przez roślinę, przez co utrzymuje się swoista homeostaza między gospodarzem i bakteriami. Zależność ta daje możliwość wykorzystania tego procesu w rolnictwie, co stanowiłoby alternatywę dla nawozów sztucznych i dało szansę na uzyskanie lepszych plonów naturalnymi metodami.
Literatura:
1. Pietr S. J., Pisarska K. (2014) ,,Bakterie endofityczne – ich pochodzenie i interakcje z roślinami” Postępy mikrobiologii 53, 2, 141–151
2. Klama J. (2004) ,,Współżycie endofitów bakteryjnych z roślinami” Acta Scientarum Polonorum Agricultura 3(1), 19-28
3. Krzesłowska M., Rabęda I., Woźny A. (2011) ,,Bakterie i grzyby mikoryzowe w fitoremediacji metali śladowych” Kosmos 60, 3-4, 423-433
4. Kubiak K., Oszako T., Siebyła M., Tkaczyk M. (2013) ,,Obecność endofitów w tkankach roślin a odporność na patogeny korzeniowe”
5. Brader G., Compant S., Mitter B., Sessitsch A.,Trognitz F. (2014) ,,Metabolic potential of endophytic bacteria” Current Opinion in Biotechnology 27, 30-37
6. Carell A. A., Frank A. C. (2014) ,,Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation” Front. Microbiol.